-
1 crossing-over
crossing-over кроссинговер (перекрёст хромосом)crossing-over кроссинговер, перекрест, обмен участками гомологичных хромосом в мейозне, обусловливающий разъединение сцепленных геновcrossing-over value величина кроссинговера, частота кроссинговера между двумя генами (стат.)complementary double crossing-over двойной комплементарный кроссинговерcompound crossing-over сложный кроссинговерdouble crossing-over двойнойdouble-strand crossing-over обмен двух нитей (двойной кроссинговер, при котором второй обмен происходит между хроматидами, участвовавшими в первом обмене)effective crossing-over эффективный кроссинговерfour-strand crossing-over обмен четырёх нитей (двойной кроссинговер, при котором второй кроссинговер происходит между хроматидами, не участвовавшими в первом)gonial crossing-over гониальный кроссинговерillegitimate crossing-over неправильный кроссинговерillegitimate crossing-over неправильный кроссинговер, кроссинговер у гаплоида или полиплоида между гомологичными и редуплицировавшими сегментами двух хромосом, различных по структуре и нормально не конъюгирующихinterallelic crossing-over межаллельный кроссинговерinterallelic crossing-over неправильный кроссинговерinversion crossing-over инверсионный кроссинговерinversion crossing-over инверсионный кроссинговер, кроссинговер между гомологичными хромосомами, отличающимися по двум инверсиям, которые имеют, однако, общий инвертированный участокprogressive crossing-over прогрессивный кроссинговерprogressive double crossing-over двойной прогрессивный кроссинговерregressive double crossing-over двойной регрессивный кроссинговерsister-strand crossing-over перекрёст сестринских нитей хромосом или хроматидsomatic crossing-over соматический кроссинговерsomatic crossing-over соматический кроссинговер, кроссинговер, происходящий при митозе в соматических клеткахthree-strand crossing-over обмен трёх нитей (двойной кроссинговер, при котором второй обмен происходит между одной из участвовавших в первом обмене хроматид и не участвовавшей в нём хроматидой)two-strand crossing-over обмен двух нитей (двойной кроссинговер, при котором второй обмен происходит между хроматидами, участвовавшими в первом обмене)unequal crossing-over неравный кроссинговерunequal crossing-over неравный кроссинговер, асимметричный кроссинговер (при котором перекрест гомологов происходит не в идентичных точках, благодаря чему одна хроматида содержит оба аллеломорфа данного гена, а другая не содержит его вовсе)English-Russian dictionary of biology and biotechnology > crossing-over
-
2 equatorial plate
Скопление хромосом в медиальной плоскости веретена во время метафазы клеточного деления; часто (у животных) хромосомы на Э.п. располагаются упорядоченно (центромерами к ее центру и хроматидами - к периферии).* * *Англо-русский толковый словарь генетических терминов > equatorial plate
-
3 reciprocal chiasmata
Две хиазмы, образуемые одними и теми же хроматидами.* * *Хиазмы реципрокные — две хиазмы, образованные одними и теми же хроматидами.Англо-русский толковый словарь генетических терминов > reciprocal chiasmata
-
4 theory
theory теорияtheory of evolution эволюционный теорияtheory of genasthenia теория функциональной валентности геновtheory of gene starvation теория голодания генаtheory of germ plasm теория зародышевой плазмыallogenetic theory аллогенетическая теорияbalance theory of sex теория полового балансаcataclysmic theory теория катастрофchiasmatype theory теория хиазматипииchiasmatype theory хиазматипическая теория, согласно которой хиазмы образуются только между сестринскими хроматидами, благодаря чему кроссинговер происходит между двумя хроматидами, а не целыми хромосомамиconversion theory теория конверсии (генов)displacement theory теория континентального дрейфа, теория Вегенераerror theory биом. теория ошибокfrozen accident theory теория замороженного случаяgene replica theory теория генных репликlateral-chain theory теория боковых цепейmass-flow theory теория потока веществmnetic theory теория мнемии, теория, которая приписывает явление наследственности скрытой памяти о прошедших генерацияхone gene - one enzyme theory теория один ген - один ферментpreformation theory теория преформизмаpreformation theory теория преформации, теория, согласно которой каждое яйцо или сперматозоид животного содержит уже в себе взрослый организм в миниатюре, для развития которого нужно только соответствующее питание и времяpresence and absence theory теория присутствия - отсутствия, связывающая доминантность с присутствием гена, а рецессивность - с его отсутствиемpresence-and-absence theory теория присутствия - отсутствияrecapitulation theory теория рекапитуляции, теория о том, что развитие организмов является кратким повторением истории их эволюцииsaltation theory сальтационная теория, теория скачковseparation theory теория изоляцииside-chain theory теория боковых цепейstereochemical theory стереохимическая теорияtarget theory биом. теория попаданий, теория удачtrace theory of memory следовая теория памятиtransition state theory теория переходного состоянияunitarian theory унитарная теория (кроветворения)English-Russian dictionary of biology and biotechnology > theory
-
5 exchange
обмен; физиол. обменfetal-maternal exchange — трансплацентарный перенос, трансплацентарный транспорт
segmental exchange — обмен сегментами ( генов), сегмент-рекомбинация
-
6 model
модельmodel of antigen-specific help — модель антиген-специфического хелперного сигнала, модель когнатной помощи
allelic deletion model — модель делеции в аллельной хромосоме, модель Хонжо-Катаоки
antigen-bridge model — модель антигенного мостика, модель Митчисона ( модель кооперативного взаимодействия Т- и В-лимфоцитов)
antigenic model — антигенная модель, ( молекулярная) модель антигена
association model — ассоциативная модель (одна из моделей активации T-клетки, основанная на ключевой роли сложного кооперативного участия CD4+, CD8+ и T-клеточных рецепторов)
attenuated vaccine model — модель аттенюированной [ослабленной] вакцины (экспериментальная модель для изучения иммунитета к повторному заражению, основанная на предварительном введении животному аттенюированной вакцины)
barrel stave model — модель «бочарных клёпок» (одна из моделей образования пор в мембране клетки-мишени путём агрегации мономерных мембранных белков в момент контакта с лимфоцитом-киллером)
B-cell model — B-клеточная модель иммунного ответа, модель гуморального иммунного ответа
Bell's model — модель двухстадийного связывания антигенов с клетками, ( математическая) модель Белла
birth-death model — модель «рождения-смерти» (модель Тилла, объясняющая механизмы дифференцировки кроветворных клеток)
bracelet model — модель «браслета» ( модель олигомерной структуры молекулы иммуноглобулина M)
Bruni's model — математическая модель Бруни, интегрально-дифференциальная равновесная модель ( гуморального иммунного ответа)
Clarke-Kirby model — модель Кларка-Кирби (одна из моделей естественной селекции вариантов гистосовместимости в системе мать-плод)
clonal deletion model — клонально-делеционная модель, гипотеза делении клонов
Cohen's model — ( математическая) модель индукции антителогенеза, модель Коэна
computer graphic model — компьютерно-графическая модель (напр. структурной организации молекулы антигена)
concomitant immunity model — модель премунитета (модель для изучения иммунитета к повторному заражению, основанная на предварительном введении экспериментальному животному минимальной дозы микробной взвеси)
Delisi-Perelson model — модель перекрёстных сшивок поверхностных иммуноглобулинов клетки с соответствующими лигандами, пэтчинг-модель, модель Делизи-Перельсона
determinant selection model — модель селекции детерминанты ( одна из моделей механизма презентации антигена макрофагами T-лимфоцитам)
hemopoietic inductive microenvironment model — модель индуцирующего влияния кроветворного микроокружения
Hoffman's model — модель Хоффмана (модель, отражающая различный характер взаимодействия между позитивными и негативными клонами иммуноцитов при разных состояниях иммунной системы)
integro-differential equation model — математическая модель Бруни, интегрально-дифференциальная равновесная модель ( гуморального иммунного ответа)
Jilek's model — ( математическая) модель кинетики гуморального иммунного ответа, модель Илека
Lefever-Garay model — математическая модель иммунологического надзора, модель Лефевра-Гарая
looping out-excision model — модель выпетливания-вырезания (модель реаранжировки иммуноглобулиновых генов при участии сайт-специфических эндонуклеаз)
looping out-excision-reintegration model — модель выпетливания-вырезания-реинтеграции ( модель интегративной реаранжировки иммуноглобулиновых генов)
mini-peptide model — минипептидная модель, модель псевдогенных, источников (гипотеза механизмов разнообразия антител, основанная на сайт-специфической рекомбинации сегментов псевдогенов и смысловых генов, кодирующих области антидетерминант)
open-ended network model — каскадная модель теории сети, модель открытой идиотипической сети, модель Рихтера
patching model — модель перекрёстных сшивок поверхностных иммуноглобулинов клетки с соответствующими лигандами, пэтчинг-модель, модель Делизи-Перельсона
peptidic self model — идиопептидная модель ( модель иммунного ответа с участием идиотипических пептидов)
platelet demand model — модель тромбоцитопоэза «по запросу» (основанная на существовании обратной связи между предшественниками и продуктами каждой стадии дифференцировки)
population dynamic model — популяционно-динамическая модель ( математическая модель индукции антителогенеза и толерантности)
predator-prey model — модель «хищника-жертвы» (математическая модель гуморального иммунного ответа на метаболизирующие антигены)
Raff-Cantor model — стадиеспецифическая модель дифференцировки и созревания T-лимфоцитов, модель Раффа-Кантора
Richter's model — каскадная модель теории сети, модель открытой идиотипической сети, модель Рихтера
sequential model of macrophage-lymphocyte interaction — гипотеза «застёжки молнии», «телеграфная» гипотеза, гипотеза Эллнера (гипотеза процессинга антигена в макрофагах и его дальнейшего представления T-лимфоцитам)
stem cell competition model — модель конкуренции стволовых клеток ( гуморальная модель регуляции гемопоэза)
stochastic model of stem cell differentiation — стохастическая модель дифференцировки стволовой клетки
Szilard's model — селекционная модель ( иммунитета), модель Сциларда
T-B cell model — модель кооперации [модель взаимодействия] T- и B-клеток
threshold models — ( математические) «пороговые» модели ( гуморального иммунного ответа на метаболизирующие антигены)
-
7 Holliday model
Модель, описывающая механизм кроссинговера между хроматидами, в соответствии с ней 2 несестринских двухцепочечных молекулы ДНК, между которыми происходит рекомбинация, выстриваются друг против друга, и в цепях одной и той же полярности в идентичных сайтах возникают одноцепочечные разрывы, каждая из расщепленных цепей спаривается с комплементарным участком нерасщепленной цепи противоположного дуплекса, что после лигирования приводит к образованию точки ветвления, которая может перемещаться вдоль цепей ДНК branch migration; при этом в каждой из рекомбинирующих молекул ДНК происходит замена сегмента цепи ДНК на цепь рекомбинирующего партнера, после изомеризации комплекса с образованием Х-образной структуры (структура Холлидея) происходит разделение молекул ДНК путем внесения эндонуклеазных разрывов и лигирования; М.Х. подтверждается данными авторадиографического анализа и электронной микроскопии.* * *Холлидея модель — модель, описывающая механизмы и последовательность событий, происходящих в процессе кроссинговера между двумя хромосомами гомологичными. Согласно Х. м., две двухцепочечные несестринские молекулы ДНК выстраиваются друг против друга и в идентичных сайтах происходят одноцепочечные разрывы (в цепочках с одинаковой полярностью). Возникшие цепочки спариваются с комплементарным участком нерасщепленной цепочки противоположного дуплекса, происходит их лигирование и образуется точка ветвления, которая может перемещаться вдоль цепей ДНК. В каждой из рекомбинируемых молекул происходит замена сегмента цепи ДНК на цепь рекомбинирующего партнера, изомеризация комплекса с созданием Х-образной структуры (структуры Холлидея) и разделение молекул ДНК вследствие эндонуклеазных разрывов и лигирования. Х. м. подтверждается данными радиоавтографии и электронной микроскопии.Англо-русский толковый словарь генетических терминов > Holliday model
-
8 meiosis
мейоз, деление созреванияДвухступенчатое деление клеток, приводящее к образованию из диплоидных клеток гаплоидных, что является основным этапом гаметогенеза; выделяют 3 типа М.: зиготный, или начальный (у многих грибов и водорослей) - происходит сразу после оплодотворения и приводит к образованию гаплоидного таллома или мицелия, гаметный, или конечный (у всех многоклеточных животных и у некоторых низших растений) - происходит в половых органах и приводит к образованию гамет, споровый, или промежуточный (у высших растений) - происходит перед цветением и приводит к образованию гаплоидного гаметофита, у простейших встречаются все 3 типа М.; М. включает два деления, разделенных интеркинезом interkinesis( но не всегда обязательным), - I деление характеризуется очень длинной, дифференцированной на стадии профазой, во II профаза и метафаза могут выпадать; удвоение ДНК происходит только перед I делением М.; однако прежняя точка зрения о том, что в I делении расходятся гомологичные хромосомы, а во II - хроматиды (т.е. I - редукционное деление, а II - эквационное) не подтверждается: в I делении расходятся либо хромосомы, либо хроматиды, а во II - наоборот; М. был открыт У.Флеммингом у животных в 1882 и Э.Страсбургером у растений в 1888.* * *Мейоз — деление клеточного ядра, предшествующее образованию половых клеток и связанное с уменьшением (редукцией) числа хромосом, свойственного соматической клетке, в 2 раза. Различают 3 типа М.:а) начальный, или зиготный, — происходит сразу после оплодотворения, приводя к образованию гаплоидного таллома или мицелия (у многих грибов и водорослей);б) конечный, или гаметный, — происходит в половых органах и приводит к образованию гамет (у всех многоклеточных животных и у некоторых низших растений);в) споровый, или промежуточный, — происходит перед цветением и приводит к образованию гаплоидного гаметофита (у высших растений).У простейших выявлены все 3 типа М. Гаметный (конечный) тип М. состоит из двух следующих одно за другим делений: I, которое включает в себя очень длинную, состоящую из нескольких стадий профазу и метафазу, и II, в котором могут выпадать профаза и метафаза. Профазы I деления подразделяется на 5 последовательных стадий: лептотену (лептонему), зиготену (зигонему), пахитену (пахинему), диплотену (диплонему) и диакинез. В течение лептотены хромосомы имеют вид тонких нитей с ясно различимыми хромомерами. Все хромосомы часто ориентированы одним или обоими концами и контактируют с одним участком ядерной мембраны, образуя конфигурацию «букета». Каждая хромосома состоит из 2 хроматид, однако это остается неразличимым до пахитены (репликация ДНК и удвоение ее диплоидного количества происходят до наступления лептотены). В диплоидных соматических клетках (2N) хромосомы присутствуют в виде N пар и каждая хромосома является репликантом одной из родительских хромосом самца и самки в данной зиготе. В ядрах соматических клеток большинства организмов гомологичные хромосомы не образуют пары. В М. в течение стадии зиготены происходит синапсис гомологичных хромосом: образование пар начинается в ряде точек и продолжается до полного завершения конъюгации (см. Конъюгация хромосом). Этот процесс сопровождается формированием синаптонемального комплекса. Когда синапсис заканчивается, реальное число хромосомных нитей равно половине того числа, которое было ранее, и они различимы в ядре как биваленты, а не единичные хромосомы. На стадии пахитены каждая парная хромосома разделяется на две сестринские хроматиды (за исключением центромеры). В результате продольного деления каждой гомологичной хромосомы на 2 хроматиды в ядре образуется N групп из 4 хроматид, лежащих параллельно друг другу, называемых тетрадами. Происходит локализованный разрыв с последующим обменом участками между несестринскими хроматидами — кроссинговер (см). Этот процесс сопровождается синтезом конститутивной ДНК в количестве меньшем, чем 1% от всего его количества в ядре. Обмен между гомологичными хромосомами приводит к образованию кроссоверных хроматид (см. Кроссоверы), содержащих генетический материал и отцовского, и материнского происхождения. На протяжении стадии диплотены одна из пар сестринских хроматид в каждой из тетрад начинает отделяться от др. пары. Однако хроматиды не разделяются в том месте, где имел место обмен, — в таких районах частичного перекрытия хроматиды образуют крестообразную структуру, называемую хиазмой. Число хиазм зависит от вида хромосомы и прямо пропорционально ее длине. Затем происходит терминализация хиазм, которая продолжается в течение диакинеза до тех пор, пока все хиазмы не достигнут концов тетрад и гомологи смогут разделиться во время анафазы. В диакинезе хромосомы плотно спирализуются, укорачиваются и утолщаются, образуя группу компактных тетрад, хорошо упакованных в ядре, чаще всего около его мембраны. Терминализация полностью завершается и исчезает ядрышко. Во время I деления исчезает оболочка ядра и тетрады располагаются в области экватора, где находится веретено деления. Хроматиды тетрад разъединяются т. обр., что происходит разделение материнского и отцовского генетического материала, за исключением дистального участка, где произошел кроссинговер. Во время I деления образуются 2 вторичных гаметоцита, которые содержат диады, окруженные ядерной оболочкой. II деление начинается после короткой интерфазы, в течение которой хромосомы не спирализуются. Ядерная мембрана исчезает, и диады располагаются на метафазной пластинке. Хроматиды каждой диады эквивалентны друг другу (за исключением дистальных участков с точками, претерпевшими кроссинговер), центромера делится, и каждая хромосома получает возможность уйти в отдельную клетку. У животных во время II деления образуются 4 сперматиды или оотиды с гаплоидным набором хромосом, окруженные ядерной мембраной. М., т. обр., обеспечивает механизм, посредством которого происходит обмен генетическим материалом между гомологичными хромосомами и каждая гамета получает по одной из пары хромосом. В последнее время появляются сообщения о том, что прежняя точка зрения о расхождении гомологичных хромосом в I делении М. (редукционное деление), а хроматид — во II (эквационное) не подтверждается: в I делении расходятся либо хромосомы, либо хроматиды, а во II — наоборот. М. открыт У.Флемингом у животных в 1882 г. и Э.Стасбургером у растений в 1888 г.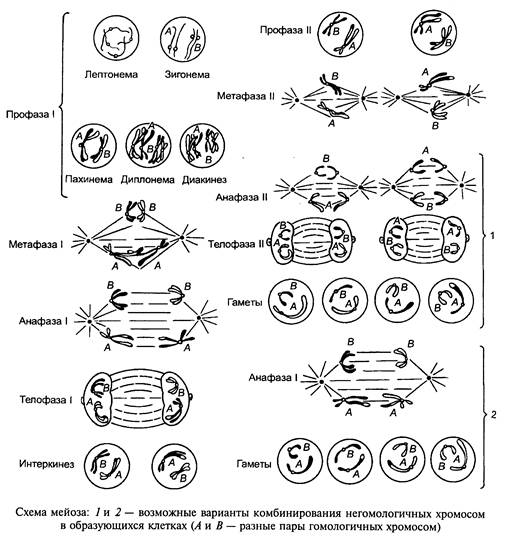
Англо-русский толковый словарь генетических терминов > meiosis
-
9 mid-body
см. equatorial plate* * *Англо-русский толковый словарь генетических терминов > mid-body
-
10 nuclear plate
см. equatorial plate* * *Англо-русский толковый словарь генетических терминов > nuclear plate
-
11 polytene chromosome
Гигантская хромосома некоторых соматических клеток, прошедших процесс политенизациии polyteny; неравномерная спирализация позволяет выделять вдоль П.х. характерный набор большого числа имеющих высокоспецифичное распределение дисков, в результате чего становится возможным обнаружение мелких хромосомных перестроек и даже определение локализации отдельных генов; П.х. известны в клетках слюнных желез, мальпигиевых сосудов, трихогенных и некоторых др. клетках насекомых (двукрылых и бессяжечниковых), в клетках растений, грибов (род Sciara), простейших (инфузории); в 1989 В.Сорса опубликовал исчерпывающую 2-томную монографию по П.х. дрозофил.* * *Политенная хромосома — гигантская «канатоподобная» хромосома, состоящая из параллельно расположенных идентичных хроматид. Хроматин в них гиперспирализован в определенных участках, и связи между хроматидами образованы вертикально по отношению к продольной оси хромосомы. П. х. обнаружены у небольшого числа организмов. Они присутствуют в макроядре водорослей р. Ciliate, в синергидах и антиподах семяпочек некоторых покрытосеменных растений и в различных тканях особей р. Dipterae (Двукрылые). Наиболее глубоко изучены П. х. слюнных желез у Drosophila (см. Бальбиани кольца. otu-мутация).Англо-русский толковый словарь генетических терминов > polytene chromosome
-
12 sister chromatid exchanges
сестринские хроматидные обмены, СХОПроцесс обмена участками между сестринскими хроматидами sister chromatids в мейозе или митозе; СХО могут быть выявлены включением аналогов оснований, маркирующих при специальном окрашивании одну из цепей двухцепочечной молекулы ДНК - например, включением 5-бромдезоксиуридина 5-bromodeoxyuridine; в норме СХО происходят достаточно часто (3-8 в разных случаях в расчете на 1 клетку у человека), однако их частота может существенно увеличиваться при действии некоторых мутагенов, т.е. определение частоты СХО является эффективным тестом на мутагенность; увеличение количества СХО сопровождает некоторые НЗЧ - например, синдром Блума Bloom syndrome.Англо-русский толковый словарь генетических терминов > sister chromatid exchanges
-
13 sister chromatid reunion
Слияние идентичных «кусков» сестринских хроматид sister chromatids, образовавшихся в результате изохроматидного разрыва редуплицированной хромосомы; С.х.о. может происходить только между центрическими фрагментами, только ацентрическими или между фрагментами обоих типов (полное С.х.о.).* * *Воссоединение сестринских хроматид — слияние идентичных, параллельно лежащих обломков сестринских хроматид. В этом же значении употребляются также термины «воссоединение сестринских нитей» и «рекомбинация между сестринскими хроматидами». Обмен между четырьмя фрагментами, образовавшимися в результате изохроматидного разрыва редуплицированной хромосомы, приводит к трем типaм В. с. х.:а) полное воссоединение, т. е. воссоединение центрических и ацентрических фрагментов сестринских хроматид (SU);б) воссоединение только ацентрических фрагментов (NUp) при отсутствии воссоединения проксимальных фрагментов;в) воссоединение только центрических фрагментов (NUd) при отсутствии воссоединения дистальных. Когда воссоединение вообще отсутствует, применяется символ NUpd, т. е. «невоссоединение» проксимальных и дистальных фрагментов. При воссоединении центрических фрагментов возникает дицентрическая хроматида, которая в случае расхождения центромер к разным полюсам растягивается между ними и обычно разрывается. Если обе центромеры отходят к одному полюсу, то разрыва дицентрической хроматиды не происходит. Ацентрические фрагменты в цитоплазме исчезают.Англо-русский толковый словарь генетических терминов > sister chromatid reunion
-
14 sister strand reunion
Слияние идентичных «кусков» сестринских хроматид sister chromatids, образовавшихся в результате изохроматидного разрыва редуплицированной хромосомы; С.х.о. может происходить только между центрическими фрагментами, только ацентрическими или между фрагментами обоих типов (полное С.х.о.).* * *Воссоединение сестринских хроматид — слияние идентичных, параллельно лежащих обломков сестринских хроматид. В этом же значении употребляются также термины «воссоединение сестринских нитей» и «рекомбинация между сестринскими хроматидами». Обмен между четырьмя фрагментами, образовавшимися в результате изохроматидного разрыва редуплицированной хромосомы, приводит к трем типaм В. с. х.:а) полное воссоединение, т. е. воссоединение центрических и ацентрических фрагментов сестринских хроматид (SU);б) воссоединение только ацентрических фрагментов (NUp) при отсутствии воссоединения проксимальных фрагментов;в) воссоединение только центрических фрагментов (NUd) при отсутствии воссоединения дистальных. Когда воссоединение вообще отсутствует, применяется символ NUpd, т. е. «невоссоединение» проксимальных и дистальных фрагментов. При воссоединении центрических фрагментов возникает дицентрическая хроматида, которая в случае расхождения центромер к разным полюсам растягивается между ними и обычно разрывается. Если обе центромеры отходят к одному полюсу, то разрыва дицентрической хроматиды не происходит. Ацентрические фрагменты в цитоплазме исчезают.Англо-русский толковый словарь генетических терминов > sister strand reunion
-
15 two strand double crossing-over
двойной двуххроматидный (прогрессивный) кроссинговер, обмен двух нитейДвойной кроссинговер, при котором второй обмен происходит между хроматидами, участвовавшими в первом обмене.Англо-русский толковый словарь генетических терминов > two strand double crossing-over
-
16 double-strand crossing-over
Макаров: обмен двух нитей (двойной кроссинговер, при котором второй обмен происходит между хроматидами, участвовавшими в первом обмене)Универсальный англо-русский словарь > double-strand crossing-over
-
17 equatorial (nuclear) plate
Общая лексика: эквато (скопление хромосом в медиальной плоскости веретена во время метафазы клеточного деления; часто (у животных) хромосомы на Э. п. располагаются упорядоченно (центромерами к её центру и хроматидами - к периферии))Универсальный англо-русский словарь > equatorial (nuclear) plate
-
18 equatorial plate
1) Общая лексика: (nuclear) эквато (скопление хромосом в медиальной плоскости веретена во время метафазы клеточного деления; часто (у животных) хромосомы на Э. п. располагаются упорядоченно (центромерами к её центру и хроматидами - к периферии))2) Медицина: экваториальная пластинка3) Генетика: ядерная пластинка4) Макаров: метафазная пластинка -
19 four-strand crossing-over
Макаров: обмен четырёх нитей (двойной кроссинговер, при котором второй кроссинговер происходит между хроматидами, не участвовавшими в первом)Универсальный англо-русский словарь > four-strand crossing-over
-
20 interchromosomal sister chromatid exchange
Иммунология: межхромосомный обмен сестринскими хроматидамиУниверсальный англо-русский словарь > interchromosomal sister chromatid exchange
См. также в других словарях:
Обмен между сестринскими хроматидами — * абмен паміж сястрынскімі храматыдамі * sister chromatid exchange обмен генетическим материалом между двумя хроматидами одно и той же хромосомы в процессе деления клетки … Генетика. Энциклопедический словарь
Обмен между несестринскими хроматидами — * абмен паміж несястрынскімі храматыдамі * nonsister chromatid exchange обмен сегментами между гомологичными партнерами в профазе I мейоза (пахитена). Разрывы в хроматидах происходят в одних и тех же местах, и обмен между определенными сегментами … Генетика. Энциклопедический словарь
Митоз — Фазы митоза Митоз (греч … Википедия
Сестринский хроматидный обмен — Схематическое изображение сестринского хроматидного обмена … Википедия
Кроссинговер двойной д перекрест — Кроссинговер двойной, д. перекрест * красінговер двайны, д. перакрыж * double crossing over совпадение, или коинциденция (см.), двух кроссинговеров в двух точках одной и той же пары гомологичных хромосом, результатом чего являются два обмена.… … Генетика. Энциклопедический словарь
Хромосомы — (от Хромо... и Сома органоиды клеточного ядра, совокупность которых определяет основные наследственные свойства клеток и организмов. Полный набор Х. в клетке, характерный для данного организма, называется Кариотипом. В любой клетке тела… … Большая советская энциклопедия
ТЕЛОФАЗА — ТЕЛОФАЗА, стадия ДЕЛЕНИЯ КЛЕТКИ, следующая после АНАФАЗЫ. Это конечная фаза МИТОЗА и МЕЙОЗА. В митозе телофаза включает деление ЦИТОПЛАЗМЫ для образования двух дочерних клеток с тем же числом ХРОМОСОМ, что и в ядре исходной клетки. Разделенные… … Научно-технический энциклопедический словарь
Диплотена (Diplotene) — четвертая стадия первой профазы мейоза, при которой между спаренными хроматидами гомологичных хромосом образуется перекрест, и затем они начинают разделяться. Источник: Медицинский словарь … Медицинские термины
МЕЙОЗ — (от греч. meiosis уменьшение), деления созревания, особый способ деления клеток, в результате к рого происходит редукция (уменьшение) числа хромосом и переход клеток из диплоидного состояния в гаплоидное; осн. звено гаметогенеза. М открыт В.… … Биологический энциклопедический словарь
РАСЩЕПЛЕНИЕ — в генетике, появление в потомстве гибрида особей (клеток) разного генотипа (Р. по генотипу) или обусловленное генотнпически различие потомков по проявлению признака (Р. по фенотипу). Закономерности Р. признаков в определённом количеств,… … Биологический энциклопедический словарь
ХРОМОСОМЫ — (от хромо... и сома), органоиды клеточного ядра, являющиеся носителями генов и определяющие наследств, свойства клеток и организмов. Способны к самовоспроизведению, обладают структурной и функциональной индивидуальностью и сохраняют её в ряду… … Биологический энциклопедический словарь